
Unlock the Cellular Power of Intense Exercise
Why Intensity Is the Ultimate Differentiator for Aging Well

A Note Before You Begin
This article makes a case that high intensity exercise benefits aging, healthspan, functional ability, and disease risk based on peer-reviewed research. It is not medical advice. If you’ve been sedentary, have any chronic health conditions, or are over forty and haven’t exercised intensely in years, consult a physician before adding high-intensity work to your routine. 1,2 Intensity is a tool (a powerful one) but it must be worked up to gradually. The goal is to build capacity over months and years, not to sprint into a heart attack next Tuesday. Start where you are. Progress deliberately. When in doubt, get cleared by someone who knows your heart, your history, and your bloodwork. 3
The Moment the Body Wakes Up
Two men pedal stationary bikes in the same gym, separated by six feet of rubber flooring and an almost infinite biological distance.
The first rides easy. Heart rate: 112. His breathing is slow, measured, almost absent-minded. He scrolls through emails on the phone propped against the console, occasionally swiping, occasionally frowning at a subject line. His legs turn with metronomic patience. If you asked him to hold a conversation, he could. If you asked him to sing, he probably could manage that too. 4-6
The second man inhabits a different universe.
He is bent over the handlebars, hands clamped white against the grips, legs driving the pedals with a rhythm that borders on violent. Heart rate: 171. His chest heaves. Sweat streams down his temples and pools in the hollows of his collarbones. If you asked him a question, you’d get three gasped words at most. 7,8
The room has narrowed to a tunnel: nothing exists but the next stroke, the next breath, the negotiation between will and the accumulating burn in his quadriceps.
Both men are fifty-two years old. Both woke early to “get their cardio in.” But deep in their muscle fibers, two entirely different biological conversations are taking place. 9-12
In the first man’s cells, the machinery hums along at idle. Energy supply keeps pace with demand. The slow-twitch muscle fibers, the endurance specialists, built for steady, low-power work, handle the load without stress.
To understand why this easy effort won’t trigger adaptation, and why the second man’s brutal pace will, you have to zoom in to the cell’s energy currency system.
ATP (adenosine triphosphate) is the cell’s spendable form of energy. Each muscle contraction is powered by snapping off a phosphate from ATP, converting it into ADP (adenosine diphosphate) and releasing usable energy. ADP is not useless. It’s a partly spent coin that can be recycled back into ATP. During lower levels of activity, this recycling occurs at a one-to-one ratio, at a rate that allow ATP levels to remain high and the cellular energy economy to stay balanced. 13-15
However, as activity picks up into intense exercise, energy demand spikes. The normal mechanisms of cellular energy production can’t keep up: ADP can’t be recycled into ATP quickly enough. 16,17 The first backup system uses phosphocreatine to donate phosphate groups directly to ADP, but it depletes within seconds of all-out effort. (This is where creatine supplementation enters the picture, but that’s a subject for a future article.) 18 When even that system can’t keep pace, a second salvage system kicks in: an enzyme called adenylate kinase that converts two ADP into one ATP, leaving behind one AMP (adenosine monophosphate). That leftover AMP is the key. Unlike ADP, AMP can’t easily be recycled. As intense activity continues, AMP accumulates. And the cell has a sensor watching for exactly this signal. 19-21

The sensor is called AMPK (AMP-activated protein kinase). 22,23 Think of it as the cell’s fuel gauge. 24 When AMP levels rise, AMPK detects the imbalance and initiates an adaptive response. 25 Some effects are immediate: shifting the cell toward fat-burning and increasing glucose uptake. But the critical long-term effect is a signal to build more mitochondria, the power plants responsible for converting food and oxygen into usable energy, so the cell will have greater energy-producing capacity next time.
26-28
AMPK is a kinase, one member of a family of enzymes that act as molecular switches. 29,30 Kinases work by attaching a phosphate group to other proteins (a process called phosphorylation), physically changing their shape and behavior. An inactive enzyme suddenly starts working. A dormant gene gets read. A protein stuck in one part of the cell relocates to another. The cell does not need to build new machinery (which takes hours); it flips switches on machinery it already has (which takes seconds). 31 Phosphorylation is the cell’s rapid-response system.
During the first man’s moderate ride, his AMP levels stay low. AMPK barely stirs. The switches stay off. No emergency, no response.
In the second man’s cells, a cascade is already underway.
AMP is accumulating. AMPK detects the imbalance and initiates its adaptive response. 32-34 But AMPK is not acting alone. Calcium floods the muscle fibers, waking other kinases that remained dormant through the first man’s easier effort. Lactate, accumulating faster than it can be cleared, triggers phosphorylation cascades across thousands of sites: flipping switches on proteins governing mitochondrial production, glucose transport, fat metabolism, and stress protection. Deep in the mitochondria themselves, a protein that regulates the splitting and merging of these energy factories receives a signal that only arrives during intense exertion. 35-38 A signal that will, over the coming hours and days, reshape the architecture of the cell’s power grid.
Same bikes. Same gym. Same Tuesday morning. One man is maintaining. The other is adapting.
This is the difference that intensity makes. It is not a matter of degree. It is a matter of kind.
How Do I Know What “Intense” Exercise Is?
The threshold into genuinely intense exercise is often approximated as somewhere between roughly 76 and 96 percent of maximum heart rate, but those numbers are only signposts. 39-43 Absent extensive physiological testing, an individual’s maximum heart rate is difficult to accurately nail down. A 2020 study published in the International Journal of Exercise Science found “poor agreement” between measured and age-predicted maximum heart rates. 44 Similarly, in 2023, Finnish researchers determined that simple age-dependent maximum heart rate models “are not accurate or universal.” 45 The Mayo Clinic explains that depending on a variety of factors, including overall fitness level and health, an individual “may have a higher or lower maximum heart rate, sometimes by as much as 15 to 20 beats per minute” than age-based estimates would indicate. 46 In other words, the numbers are estimates. The physiology is not.
What defines intensity is not a percentage on a fitness wearable but the unmistakable physiological state it reflects. 47 The American Heart Association explains that rather than relying on heart rate alone, a person’s subjective perception of exertion is often a more reliable indicator of true fatigue. 48,49
The simplest method is the talk test, which has predicted training intensity with surprising accuracy for nearly a century: 50-54
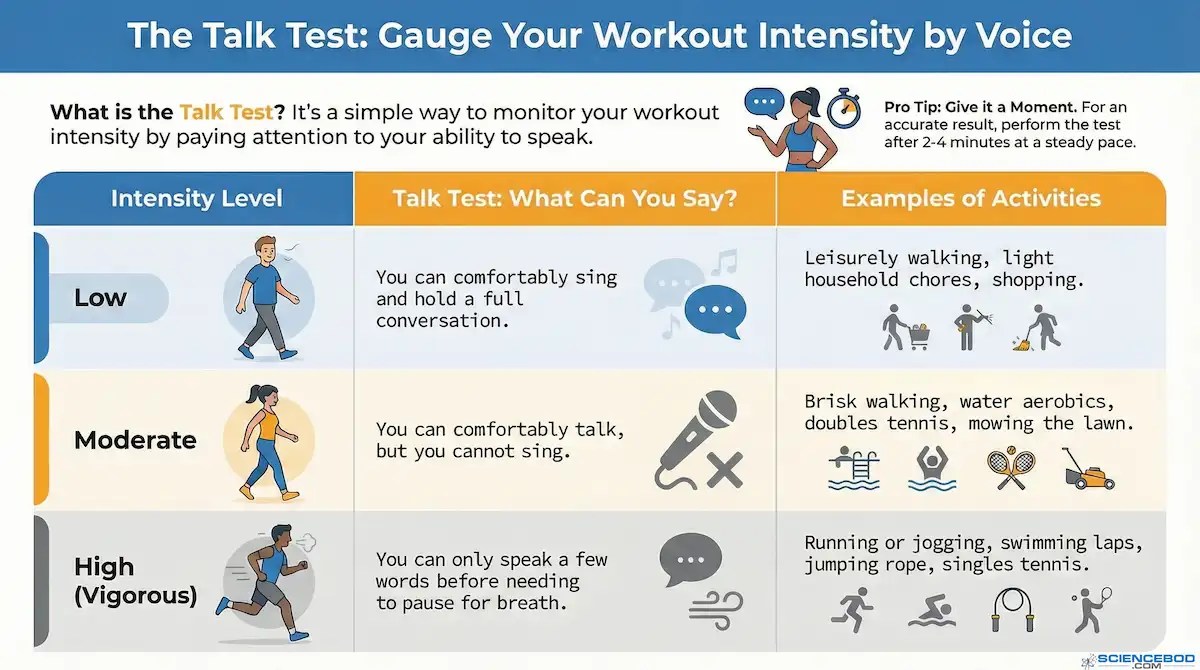
That third row is the territory that matters. Breathing becomes rapid, forceful, and dominates attention. Effort stops being something you can half-ignore while thinking about something else. Continuing requires conscious, focused commitment.
Physiologists call this border the lactate threshold: the point at which lactate accumulates in the blood faster than the body can clear it. 55,56 The ventilatory threshold, where breathing shifts from manageable to a domination of your attention, tracks it closely, which is why the “talk test” works. 57 If you can speak only in gasped fragments of three or four words, you have crossed the line. 58
Below this line, the body manages the load. Above it, the body is forced to adapt.
The Ratio That Misled Us
For decades, exercise science told us a convenient story: work harder and you’ll burn more calories. The underlying assumption was simple. What matters is energy expenditure, and two activities that burn the same number of calories should deliver roughly the same benefit. 59
Public health agencies codified this into a tidy exchange rate based on METs (Metabolic Equivalents of Task) a measure of how much energy an activity burns relative to sitting quietly. One MET equals the energy you expend at rest. 60,61 Walking briskly is about 3.5 METs. Jogging is around 7. Sprinting can hit 15 or higher. 62,63 Vigorous exercise burns about twice the calories per minute as moderate exercise. Therefore, one minute of vigorous effort should equal two minutes of moderate work. 64,65 The U.S. Centers for Disease Control states it plainly: “A rule of thumb is that one minute of vigorous-intensity activity is about the same as two minutes of moderate-intensity activity.” 66 The World Health Organization built its global recommendations on the same math: 150 minutes of moderate activity or 75 minutes of vigorous activity weekly. 67
There were two problems with this logic.
First, the ratio was based largely on self-reported data. People remembered how much they exercised, or thought they did, and researchers estimated from those fuzzy recollections. 68,69
Second, and more fundamental: calories burned is not the same as health gained. The 2:1 ratio treated exercise as a simple furnace: a way to burn fuel. But the body is not a furnace. It is an adaptive system that responds to signals, and intensity sends signals that have nothing to do with how many calories you expend. 70,71 The molecular cascades triggered by hard effort, the kinases that wake up, the mitochondrial remodeling that begins, the myokines (signaling proteins released by working muscles that travel through the bloodstream to affect distant organs) flooding the circulation, these don’t scale linearly with energy burned. They require a threshold to be crossed. 72
Wearable devices revealed just how wrong the old math was.
When researchers at the University of Sydney strapped accelerometers to over 73,000 adults and tracked them for eight years, they didn’t measure calories. They measured what actually matters: who got sick, who stayed healthy, and who died. For all-cause mortality, one minute of vigorous effort proved equivalent to 4.1 minutes of moderate activity: double the official estimate. For cardiovascular protection, the multiplier climbed to nearly 8-to-1. For type 2 diabetes prevention, 9.4-to-1. 73
“Our findings,” the researchers wrote, “are in stark contrast to the widely used current convention of a 1:2 ratio between vigorous and moderate intensity, which was derived from self-reported data.” 74
Intense exercise is not just marginally better. It is a fundamentally different physiological signal.
Vanity
As we age into our thirties (and for some with healthier lifestyles, our forties), the body becomes less forgiving of misdirected effort. 75,76 The runway is shorter. You need every workout to count.
But count toward what, exactly?
Nearly all of us say we want longer life and better healthspan. Still, honesty requires admitting that vanity is rarely absent from our motivation, and it often sits closer to the center than we would like to admit. Hungry for the appearance of youth and vitality, many go under the knife, hoping that visible youth might substitute for biological youth. Sometimes the attempt fails outright, leaving behind the stiff, embalmed look we all recognize. A face smoothed past the point of plausibility. A kind of mannequin, preserved but not really alive. And even in the best cases, where the result is subtle and technically successful, nothing underneath has changed. The cells do not repair themselves more efficiently. Mitochondria do not multiply more vigorously. Inflammation is not subdued. The biology continues aging on schedule, indifferent to the surface edit. It is not rejuvenation. It is a fresh coat of paint on a deteriorating structure. 77-79
Fortunately, we have a more effective intervention available than plastic surgery.
The research clearly demonstrates that lifestyle choices like clean diet, proper sleep, cutting out smoking and drinking, and intense exercise can make us look better as we age and healthier at the same time. A landmark identical twin study found that the sibling who looked older typically died first, leading scientists to conclude that “mortality is written on the face.” 80 Investigators from the Buck Institute for Research on Aging determined that facial photos predict blood levels of inflammatory proteins linked with aging. 81 Looking young is not merely cosmetic. It is a visible signal of biological age.
And biological age is not an abstraction. Multiple peer-reviewed studies have established that it predicts health, functional decline, disease risk, and mortality in ways that chronological age cannot. 82-86 Biological age, then, can act as a proxy for health. Given that visible appearance of youth is an indicator of biological age, all else being equal, if we look younger, we are healthier. If we look older, we are less healthy.
Looking younger encompasses more than smooth skin: subcutaneous fat distribution, bone architecture, the visible difference between strength and frailty. 87-89 These are not cosmetic details layered over health. They reflect the same underlying biology. And that biology responds to intense exercise.

Researchers at McMaster University discovered that exercise rejuvenates aging skin through a signaling protein called IL-15 (interleukin-15), released from working muscles and carried through the bloodstream to the skin. 90 There, IL-15 stimulates mitochondrial production in skin cells and boosts collagen synthesis. The key: IL-15 expression is linked to AMPK, the cellular fuel gauge that activates most robustly when intensity crosses the threshold. 91 Leisurely walks will not flip that switch. One study found that time spent in moderate-to-vigorous activity correlated with better skin elasticity and structure, while time spent in light activity showed no relationship at all. 92
If you want to look younger for longer, the prescription is the same: you have to push hard enough to trigger the cascade. The treadmill stroll that lets you scroll your phone is not rebuilding your collagen. The effort that leaves you breathless is.
What Intensity Unlocks
When researchers at Australian Catholic University used mass spectrometry to map every molecular switch that flips during exercise, they found something striking: of the more than 1,000 phosphorylation sites that changed during moderate and intense exercise, 348 responded only to high-intensity effort. During moderate exercise, they stayed completely silent. 93
Your cells contain thousands of these switches. Intense exercise flips hundreds that moderate effort leaves untouched. 94,95
Many of those switches control mitochondria. Mitochondria are not static. They form dynamic networks that constantly split apart and fuse back together. 96,97 In youth, these networks are dense, efficient, humming with electrical potential. With age and inactivity, they fragment. The networks break down into scattered islands. Energy production declines. Metabolic noise increases. The cell begins to run on the equivalent of backup generators. 98
High-intensity exercise reverses this decay. 99
In a landmark 2017 study, Robinson and colleagues put adults aged 65-80 through twelve weeks of high-intensity interval training. The results defied expectations: mitochondrial capacity increased by 69%, restoring cellular energy production to levels typically seen in people decades younger. 100 A 2023 study comparing HIIT to moderate continuous training confirmed the pattern: high-intensity work significantly outperformed for activating the master regulators of mitochondrial production. 101
Moderate exercise improves endurance and stabilizes existing cellular machinery, whereas high-intensity exercise drives deeper remodeling: reorganizing mitochondrial networks, accelerating the removal of damaged components, and building a more integrated structural framework. 102-105
The same principle applies to cellular aging. With age, tissues accumulate senescent cells. These cells no longer divide, but they do not disappear. Like a bad apple spoiling the bunch, they release inflammatory signals that degrade the surrounding environment and accelerate decline. 106,107
Hard exercise does not need to hunt these cells down to matter. What it appears to do, when the effort is intense enough, is change the environment that lets them cause damage. Skeletal muscle under real strain becomes a signaling organ. It releases myokines. When muscles work hard, they send instructions that tell cells throughout the body to clean up, repair, and stabilize.108,109
One of these signals is irisin. Irisin is released when muscle fibers burn fuel fast enough to activate a master control system known as PGC-1α. You can think of this system as a foreman in charge of the cell’s power plants and maintenance crews. When it is switched on, cells invest less in emergency responses and more in upkeep. Damaged parts are recycled. Power systems run cleaner. The conditions that let one bad apple spoil the crate are weakened. 110-113
The clearest evidence for this reset comes from stress signals we can measure. In aging muscle, the internal alarm system is stuck on. A 2025 study found that older adults carry persistently elevated markers of cellular distress. In young muscle, these signals appear briefly under stress and then shut off. With age, they do not shut off. 114
Twelve weeks of high-intensity interval training flipped that switch back. After the training, those stress markers fell to levels seen in young adults. The muscle stopped signaling constant emergency. 115
Moderate exercise helped these older adults stay functional. It slowed the slide. It did not reset the system. Only high-intensity training did.
Fertilizer for the Aging Brain
The same signal that remodels muscle and mitochondria also reaches the brain.
In a large analysis from the UK Biobank, one of the world’s largest long-term health studies following hundreds of thousands of adults, researchers used wearable motion sensors to track how much people actually moved in daily life. As little as thirty-five minutes per week of moderate to vigorous activity was associated with a forty-one percent lower risk of developing dementia. That is five minutes a day. The association held even in people classified as frail. Frailty raised baseline risk, but it did not cancel the benefit of effort. 116
This matters because it establishes a floor, not a ceiling. Minimal movement helps. But the biology that follows explains why more intense effort does more.
The mechanism most often implicated in how intense exercise protects the aging brain is a protein called Brain-Derived Neurotrophic Factor, or BDNF. When BDNF rises, brain cells form new connections more easily. Existing connections grow stronger. Circuits responsible for memory, judgment, and self-control remain functional longer. 117
BDNF does not respond equally to all movement. Exercise raises it, but intensity determines the size of the signal. In controlled human studies, higher-intensity exercise produces substantially larger increases in circulating BDNF than steady, moderate activity. Sprint interval training produces the strongest response. 118 The rule is simple.
The brain adapts most when the signal is strong enough to demand change.
High-intensity interval training appears to be especially important for executive function. These are the skills that allow planning, focus, task switching, and impulse control. They are not abstract abilities. They are the skills that keep people independent. A recent meta-analysis of randomized trials found that high-intensity interval training significantly improves executive function, along with information processing and memory. 119
These abilities decline earlier than most people think. Reviews of lifespan cognition show that processing speed and higher-order control begin to drift downward as early as the third decade of life. The change is subtle at first. Then it accumulates. 120 This is why cognitive decline feels sudden when it finally becomes obvious. The damage has been building quietly for years.
That reality carries an implication that is easy to miss. If intense exercise is one of the signals that preserves and rebuilds these circuits, then it is not something to add only when decline becomes obvious. It is something to maintain regularly across adulthood, while the brain still has capacity to protect. Waiting until symptoms appear is like reinforcing a structure only after the beams have already begun to rot.
One reason intensity matters is the metabolic signal it sends to the brain. During vigorous exercise, lactate rises sharply. Long treated as waste, lactate is now understood to be a signaling molecule. It crosses into the brain and influences pathways tied to learning and adaptation. 121,122 With sufficient metabolic strain, ketones rise as well. One of them, beta-hydroxybutyrate, increases when energy demand outpaces easy fuel supply, a state reached during sustained or high-intensity work. In animal models, this ketone has been shown to induce BDNF expression in the hippocampus. 123
The hippocampus matters because it is where memories are formed and where we learn to orient ourselves in the world. When it weakens, people get lost. Names slip. Context fades.
We know the hippocampus responds to demand because we have watched it change.
In London, taxi drivers must pass a test known as “The Knowledge.” To earn a license, they spend years memorizing every street, landmark, hotel, restaurant, and alley within several miles of Charing Cross railway station, the city’s traditional reference point. There are no shortcuts. They learn the city by repetition and correction.
Brain scans later showed that these drivers developed larger and more structurally robust hippocampi than the general population. The change was not genetic. It was earned one memorized street and landmark at a time. 124-126
The taxi drivers sent their demand signal through years of memorization. Intense exercise sends a similar signal through a different channel: the lactate and ketones that flood the brain when effort crosses the threshold. Both tell the hippocampus that more is needed. Both activate neuroplasticity.
The Physical Bank Account
The ancient Greeks understood the difference between lifespan and healthspan, even if they lacked the vocabulary. In myth, the immortal goddess Eos fell in love with the mortal Tithonus and begged Zeus to grant him eternal life. Zeus complied. But Eos had forgotten to ask also for eternal youth. Tithonus aged without dying, shriveling into a creature so withered and pitiful that the gods eventually turned him into a cicada, chirping endlessly in the corner of Eos’s palace. More years without more vitality is not a gift. It is a curse with a pulse.
So what separates those who age like Tithonus from those who remain capable?
Researchers have found a deceptively simple test that predicts which fate awaits you: grip strength. Squeeze a dynamometer, and you have just taken a snapshot of your physiological future. Studies now describe handgrip strength as a “central biomarker of healthy aging and a powerful predictor of future morbidity and mortality.” 127
At first glance, this seems absurd. How can one squeeze of the hand forecast whether you will still be alive, and functional, in the years to come?
The answer shows up most clearly in outcomes. In 2015, investigators from the Prospective Urban Rural Epidemiology study, known as PURE, followed more than 130,000 adults across seventeen countries. Lower grip strength predicted earlier death from all causes and from cardiovascular disease. It proved a stronger predictor than systolic blood pressure, one of medicine’s most relied-upon risk markers. 128
Grip strength works as a predictor because skeletal muscle is not just structural tissue. It is an endocrine system. When muscle contracts, it sends signals through the bloodstream that regulate metabolism, inflammation, and repair throughout the body. 129 Grip strength acts as a proxy for the health of this entire signaling system.
Some of the best-studied myokines illustrate why this matters. Interleukin-6 is released by working muscle and, in this context, helps improve insulin sensitivity and suppress chronic inflammation. 130 Interleukin-15 plays a role in maintaining skin structure, regulating body fat, and supporting immune cells involved in surveillance and repair.⁶ Irisin, already discussed, reflects the same principle. When effort is hard enough, muscle signals shift the body toward repair, efficiency, and long-term resilience. 131
When muscle weakens through disuse or age, this chemical conversation grows quieter. The body loses a regulatory voice it relies on to keep metabolism, immunity, and repair in balance.
The connection runs deeper still. Low handgrip strength is associated with elevated levels of C-reactive protein and interleukin-6, markers of chronic, low-grade inflammation. Researchers refer to this state as inflammaging. 132 It forms a loop. Physical inactivity allows inflammatory signals to rise. Inflammation accelerates muscle loss. Muscle loss further amplifies inflammation. The process feeds itself.
Perhaps most striking, grip strength is linked not only to functional age but to biological age at the cellular level. Studies examining DNA methylation patterns, chemical tags that help regulate which genes are active, change in highly predictable ways as the body ages and form the basis of modern measures of biological age, show that weaker grip strength is associated with accelerated epigenetic aging. 133 People with lower strength are aging faster at the molecular level, their biological clocks advancing ahead of their calendars.
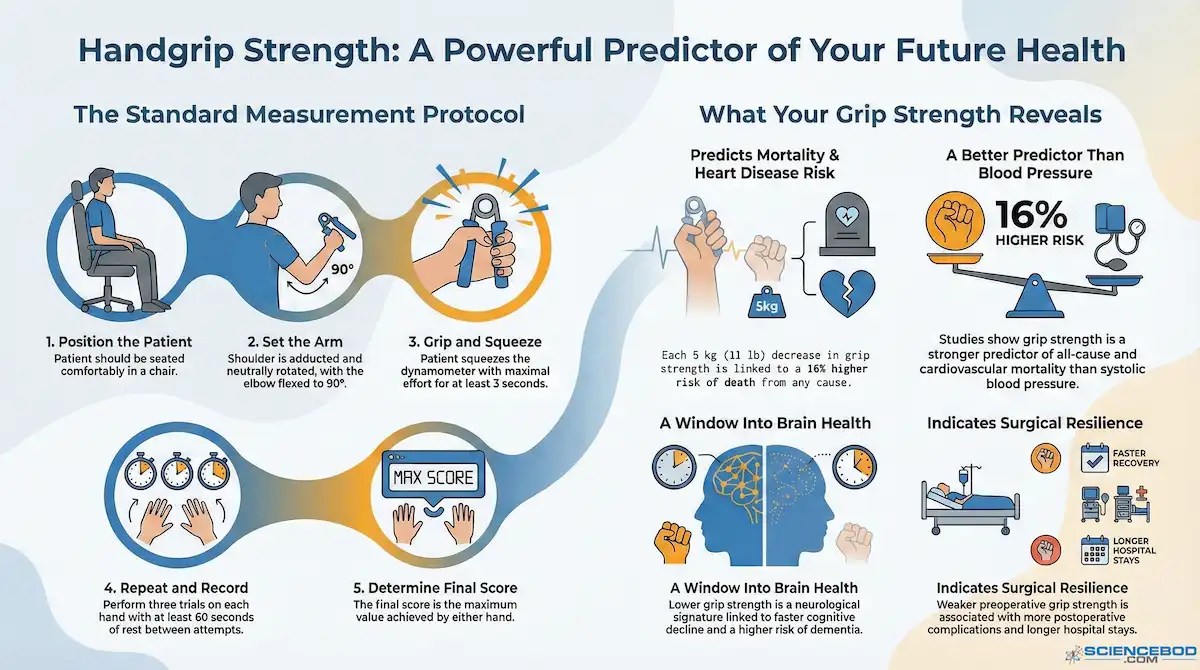
What a dynamometer measures, then, is not merely the strength of the forearm. It reflects the condition of a whole-body signaling system that governs metabolism, moderates inflammation, and influences how quickly damage accumulates inside cells.
Decline is not inevitable. Progressive resistance training improves grip strength in older adults. 134 Evidence from long-term frailty research shows that light activity often fails to change the trajectory of decline. More demanding effort is required to slow, and in some cases reverse, the slide toward dependency. 135
Bone tells the same story. Osteoblasts, the cells responsible for building bone, respond to mechanical load rather than gentle repetition. Without high-force signals, bone density erodes and fracture risk rises. 136
A hip fracture after seventy is not a minor injury. It is often the beginning of the end. One-year mortality after hip fracture commonly approaches one in four. 137 Outcomes are worse in institutionalized adults and in those with cognitive impairment. Timing matters. When surgery is delayed beyond twenty-four hours, short-term mortality and complications increase. 138 The bone breaks, and a countdown begins.
Think of physical capacity as a savings account. Moderate activity slows the withdrawals. Intense, strength-building work makes deposits. The question is whether the balance will be there when you need to draw on it.
The Molecule That Changes Everything
The same intensity threshold that builds physical reserve also triggers the body’s most powerful regenerative hormone.
In 2019, a small but provocative human trial set out to test whether that hormone could do more than preserve function. The study, called TRIIM, short for Thymus Regeneration, Immunorestoration, and Insulin Mitigation, was designed by Dr. Greg Fahy along with collaborators from Stanford and UCLA to examine a specific and controversial idea: that “thymic involution” is not merely a feature of aging, but one of its drivers. 139,140
Thymic involution is the slow shutdown of the thymus as an immune training organ. Beginning early in life, the gland steadily shrinks and its active tissue is progressively replaced by fat. By older adulthood, only a small fraction of its youthful capacity remains. 141,142
The thymus is the dojo-like “master gland” of the immune system where T cells are trained. T cells are born in the bone marrow, but they must pass through the thymus to learn how to recognize threats without attacking the body itself. As the thymus withers, the immune system loses its ability to produce fresh, “naïve” T cells. The range of threats it can recognize shrinks. Vulnerability rises. 143,144
This loss of immune training capacity is a central feature of immunosenescence, the age-related decline in immune function. As thymic output falls, the immune system becomes increasingly dependent on older, memory-based cells that are effective against familiar pathogens but poorly equipped to recognize new infections or emerging cancer cells. At the same time, dysfunctional immune cells accumulate and contribute to chronic, low-grade inflammation. 145,146
What follows is a predictable pattern. Infections become more severe. Vaccine responses weaken. The immune system becomes less effective at catching and eliminating early cancers. As the immune system stops producing fresh T cells, vulnerability to infections and cancers increases, especially later in life.147
The immune findings in TRIIM were encouraging. Imaging showed partial replacement of fatty thymic tissue with functional tissue in most participants, alongside shifts in immune cell populations consistent with renewed thymic output. 148 But another result drew far wider attention.
Using four independent DNA methylation clocks, tools that estimate biological age from the arrangement of methyl markers on the rungs of the DNA double helix, researchers measured biological age before and after treatment. After one year, participants were biologically younger than at baseline despite having aged chronologically. When adjusted for the passage of time, the net reversal averaged approximately two and a half years. One of the clocks, “GrimAge,” showed improvements that persisted six months after treatment ended.
149

This marked the first time any intervention had demonstrated measurable reversal of human epigenetic aging in living adults.
The study had clear limitations. It was small. It lacked a placebo group. Replication was required. To address those issues, Fahy’s team launched a larger follow-up trial, TRIIM-X, enrolling approximately eighty-five men and women between the ages of forty and eighty. That study is now underway and is designed to test reproducibility, expand demographics, and further evaluate safety and durability, with results expected in 2026. 150,151
Still, one detail from the TRIIM studies matters for everything discussed so far.
The TRIIM protocol used five components. Three of them, zinc, vitamin D, and DHEA, are inexpensive and available without a prescription. Metformin is also cheap, but requires a doctor willing to prescribe it off-label for longevity rather than diabetes. The truly limiting factor is human growth hormone. It is expensive. It requires a prescription. It requires medical supervision. 152
But the body already produces it.
Growth hormone is released by the pituitary gland in pulses, with the largest surge occurring during deep sleep. Exercise also stimulates its release, and the magnitude of that response depends on one variable above all others: intensity. Growth hormone secretion rises sharply once exercise crosses the lactate threshold, the same physiological border that separates moderate effort from truly intense work. 153,154
When lactate accumulates during high-intensity exercise, growth hormone levels rise dramatically. In conditions where lactate production is impaired, such as McArdle disease, this growth hormone response is often absent, underscoring the importance of crossing that metabolic threshold. 155,156
To generate a robust growth hormone surge, exercise must be sustained above this threshold. Resistance training that emphasizes metabolic stress produces particularly strong responses. The pattern holds across exercise modalities. What matters is not the activity itself, but whether the signal is strong enough to force adaptation. 157,158
This does not mean exercise replicates a pharmaceutical protocol. Endogenous growth hormone release is brief and pulsatile. The TRIIM intervention combined multiple agents, and it remains uncertain which components were most responsible for the epigenetic effects observed.
But the implication is difficult to ignore.
Intense exercise crosses the lactate threshold. Crossing that threshold triggers growth hormone release. Growth hormone was the central and most costly component of the only intervention yet shown to reverse human epigenetic aging.
The Dose, the Limits, and the Prescription
Intensity is powerful. Power requires precision. 159
For most people, better cardiorespiratory fitness protects the heart. Large studies following thousands of people over time show that improving fitness is associated with a lower risk of atrial fibrillation, the most common type of chronic irregular heartbeat and a major risk factor for stroke. The relationship is dose-dependent. Each incremental improvement in fitness correlates with a measurable drop in risk. 160
But the relationship is not linear at the extremes. In men who pile up very high volumes of endurance exercise over many years, the protective curve bends backward. Reviews and large cohort studies describe a U-shaped or J-shaped relationship: moderate and moderately high levels of exercise are protective, but chronic high-volume endurance training is associated with an increased risk of atrial fibrillation compared with non-athletes. 161-163
This is not a warning against intensity. It is a warning against relentless volume without rest. The strongest predictor of exercise-associated atrial fibrillation is not how hard any single session is, but cumulative lifetime exposure to high-volume endurance training. 164,165 Importantly, this risk elevation appears to be sex-specific, with far stronger evidence in men than in women. 166,167
The same principle governs overtraining. Overtraining syndrome is not a failure of willpower. It is a state of breakdown caused by repeated training stress without adequate recovery. Athletes with overtraining syndrome experience persistent performance decline, fatigue, mood disturbance, sleep disruption, and immune dysfunction that can last months. 168,169
At the molecular level, overtraining involves chronic inflammation and disruption of the hypothalamic–pituitary–adrenal axis, the body’s central stress-response system Instead of adapting efficiently to hard effort, the system becomes blunted and unstable. Recovery stalls. Inflammation lingers. 170-172
The most consistent solution observed in both elite performance and experimental studies is polarized training. In this model, most training time is spent at genuinely low intensity, with a smaller, deliberate fraction reserved for truly hard efforts. Observational analyses of elite endurance athletes reveal a strikingly consistent pattern, with roughly 80 percent of training volume performed at low intensity and about 20 percent at moderate-to-high intensity. 173,174
Randomized controlled trials confirm the advantage. Compared with threshold-focused or high-volume low-intensity models, polarized training produces greater improvements in maximal oxygen uptake, peak power, and endurance performance, while reducing markers of accumulated stress. 175,176
For healthspan rather than elite performance, the foundation remains straightforward. International guidelines recommend that adults accumulate 150 to 300 minutes per week of moderate-intensity activity, or 75 to 150 minutes per week of vigorous activity, or an equivalent combination. Meeting this baseline is associated with substantial reductions in cardiovascular and all-cause mortality. 177,178
Then intensity is layered on top.
For most adults, two to three sessions per week of genuine high-intensity work are sufficient. These sessions need not be long. Protocols that produce robust mitochondrial, hormonal, and molecular adaptations typically involve twenty to twenty-five minutes of focused work, including recovery periods. 179,180
The goal is not daily exhaustion. It is to deliver a signal strong enough to force adaptation, then allow time for recovery. When stress and recovery are properly balanced, intensity becomes medicine rather than poison.
Reclaim Your Biological Capital
Somewhere in your physiology, sensors are waiting for a signal worth responding to. Mitochondria are waiting to rebuild. Neurons are waiting for the surge that strengthens their connections. The pituitary gland is waiting to release a pulse of growth hormone in response to genuine effort.
The question is whether that signal will arrive.
The first man on the bike is maintaining. Perhaps that is enough for him. Perhaps he has accepted the slow erosion of capacity and the quiet dimming of reserve.
The second man is adapting. He is not just burning calories. He is sending instructions. Instructions that travel from muscle to mitochondria, to immune cells, to neurons. Instructions that say: this was hard. Prepare for more.
The choice is not between exercise and rest. It is between preservation and investment.
Stop settling for maintenance.
Reclaim your biological capital through intelligent intensity.
What comes next
The TRIIM-X trial is expected to publish results in 2026. If a larger, controlled study confirms that this protocol can reverse epigenetic aging in a broader population, it will reshape how we think about longevity interventions.
ScienceBod will be covering those results when they emerge.
In the meantime, the evidence already suggests that one component of that protocol, the growth hormone surge that arrives when you push past the lactate threshold, is available to anyone willing to earn it.
The Receipts
1 Barry A Franklin et al., “Exercise-related acute cardiovascular events and potential deleterious adaptations following long-term exercise training: placing the risks into perspective–an update: a scientific statement from the American Heart Association,” Circulation 141, no. 13 (2020).
2 “Exercise intensity: How to measure it,” Mayo Clinic Staff, Healthy Lifestyle: Fitness, Mayo Clinic, 2023, accessed December 26, 2025, https://www.mayoclinic.org/healthy-lifestyle/fitness/in-depth/exercise-intensity/art-20046887.
3 Franklin et al., “Exercise-related acute cardiovascular events and potential deleterious adaptations following long-term exercise training: placing the risks into perspective–an update: a scientific statement from the American Heart Association.”
4 Mayo Clinic Staff, “Exercise intensity: How to measure it.”
5 Alexandros Kasiakogias and Sanjay Sharma, “Exercise: The ultimate treatment to all ailments?,” Clinical Cardiology 43, no. 8 (2020), https://doi.org/https://doi.org/10.1002/clc.23369, https://onlinelibrary.wiley.com/doi/abs/10.1002/clc.23369.
6 “Good News: Short Bursts of Exercise Can Keep You Healthy,” People Staff, People.com, People Magazine, 2021, https://people.com/health/good-news-short-bursts-of-exercise-can-keep-you-healthy/.
7 Kasiakogias and Sharma, “Exercise: The ultimate treatment to all ailments?.”
8 Jose A. Rodríguez-Marroyo et al., “Relationship Between the Talk Test and Ventilatory Thresholds in Well-Trained Cyclists,” The Journal of Strength & Conditioning Research 27, no. 7 (2013), https://doi.org/10.1519/JSC.0b013e3182736af3, https://journals.lww.com/nsca-jscr/fulltext/2013/07000/relationship_between_the_talk_test_and_ventilatory.24.aspx.
9 Nolan J Hoffman et al., “Phosphoproteomics uncovers exercise intensity-specific skeletal muscle signaling networks underlying high-intensity interval training in healthy male participants,” Sports Medicine (2025).
10 Ronnie Blazev et al., “Phosphoproteomics of three exercise modalities identifies canonical signaling and C18ORF25 as an AMPK substrate regulating skeletal muscle function,” Cell Metabolism 34, no. 10 (2022).
11 Luke Olsen et al., “Exercise intensity modulates the human plasma secretome and interorgan communication,” bioRxiv (2025).
12 Ali Khalilimeybodi et al., “Systems modeling of mitochondrial dynamics in different exercise regimes,” bioRxiv (2025).
13 VW Rodwell et al., “Harper’s Illustrated Biochemistry, McGraw-Hill Education,” (2015).
14 Richard A Howlett et al., “Regulation of skeletal muscle glycogen phosphorylase and PDH at varying exercise power outputs,” American Journal of Physiology-Regulatory, Integrative and Comparative Physiology 275, no. 2 (1998).
15 James T Stull, Kristine E Kamm, and Rene Vandenboom, “Myosin light chain kinase and the role of myosin light chain phosphorylation in skeletal muscle,” Archives of biochemistry and biophysics 510, no. 2 (2011).
16 Khalilimeybodi et al., “Systems modeling of mitochondrial dynamics in different exercise regimes.”
17 Ferenc Torma et al., “High intensity interval training and molecular adaptive response of skeletal muscle,” Sports Medicine and Health Science 1, no. 1 (2019).
18 Rodwell et al., “Harper’s Illustrated Biochemistry, McGraw-Hill Education.”
19 Shalene E Hardman et al., “The effects of age and muscle contraction on AMPK activity and heterotrimer composition,” Experimental gerontology 55 (2014).
20 Torma et al., “High intensity interval training and molecular adaptive response of skeletal muscle.”
21 Rodwell et al., “Harper’s Illustrated Biochemistry, McGraw-Hill Education.”
22 Hardman et al., “The effects of age and muscle contraction on AMPK activity and heterotrimer composition.”
23 Simran Mahatme et al., “Impact of high-intensity interval training on cardio-metabolic health outcomes and mitochondrial function in older adults: a review,” Medicine and Pharmacy Reports 95, no. 2 (2022).
24 Greg Schimmack, Ralph A DeFronzo, and Nicolas Musi, “AMP-activated protein kinase: role in metabolism and therapeutic implications,” Diabetes, Obesity and Metabolism 8, no. 6 (2006).
25 Hossein Pirani et al., “Beneficial mitochondrial biogenesis in gastrocnemius muscle promoted by high-intensity interval training in elderly female rats,” Cell Journal (Yakhteh) 25, no. 1 (2022).
26 Martin J MacInnis and Martin J Gibala, “Physiological adaptations to interval training and the role of exercise intensity,” The Journal of physiology 595, no. 9 (2017).
27 Torma et al., “High intensity interval training and molecular adaptive response of skeletal muscle.”
28 Schimmack, DeFronzo, and Musi, “AMP-activated protein kinase: role in metabolism and therapeutic implications.”
29 Alexandr P Kornev, Susan S Taylor, and Hiruy S Meharena, “Evolution of a dynamic molecular switch,” IUBMB Life 71, no. 6 (2019).
30 José M López, “Digital kinases: A cell model for sensing, integrating and making choices,” Communicative & integrative biology 3, no. 2 (2010).
31 Jane B Reece et al., Campbell Biology, vol. 9 (Pearson Boston, 2014).
32 Scott K Powers et al., “Health benefits of physical activity: What role does skeletal muscle-organ crosstalk play?,” Sports Medicine and Health Science (2025).
33 Khalilimeybodi et al., “Systems modeling of mitochondrial dynamics in different exercise regimes.”
34 Hardman et al., “The effects of age and muscle contraction on AMPK activity and heterotrimer composition.”
35 Heming Chen et al., “Kinase–Phosphatase Balance in Exercise Adaptation: Phosphorylation Programs, PTM Crosstalk, and Actionable Gaps,” Frontiers in Sports and Active Living 7 (2025).
36 Hoffman et al., “Phosphoproteomics uncovers exercise intensity-specific skeletal muscle signaling networks underlying high-intensity interval training in healthy male participants.”
37 Stull, Kamm, and Vandenboom, “Myosin light chain kinase and the role of myosin light chain phosphorylation in skeletal muscle.”
38 Torma et al., “High intensity interval training and molecular adaptive response of skeletal muscle.”
39 Micah Zuhl, “Tips for Monitoring Aerobic Exercise Intensity,” (2020: American College of Sports Medicine, 2020). Exercise-intensity-infographic-PDF.pdf.
40 Carol Ewing Garber et al., “Quantity and quality of exercise for developing and maintaining cardiorespiratory, musculoskeletal, and neuromotor fitness in apparently healthy adults: guidance for prescribing exercise,” (2011).
41 Aaron J. Coutts et al., “Heart rate and blood lactate correlates of perceived exertion during small-sided soccer games,” Journal of Science and Medicine in Sport 12, no. 1 (2009/01/01/ 2009), https://doi.org/https://doi.org/10.1016/j.jsams.2007.08.005, https://www.sciencedirect.com/science/article/pii/S1440244007001934.
42 Brian R MacIntosh et al., “What is moderate to vigorous exercise intensity?,” Frontiers in physiology 12 (2021).
43 Benedikt Meixner et al., “Zone 2 intensity: a critical comparison of individual variability in different submaximal exercise intensity boundaries,” Translational Sports Medicine 2025, no. 1 (2025).
44 Mike Toczko et al., “Accuracy Of Commonly Used Age Predicted Maximal Heart Rate Equations Differs Across Levels Of Fitness: 2137,” Medicine & Science in Sports & Exercise 54, no. 9S (2022).
45 Matias Kanniainen et al., “Estimation of physiological exercise thresholds based on dynamical correlation properties of heart rate variability,” Frontiers in physiology 14 (2023).
46 Mayo Clinic Staff, “Exercise intensity: How to measure it.”
47 Franklin et al., “Exercise-related acute cardiovascular events and potential deleterious adaptations following long-term exercise training: placing the risks into perspective–an update: a scientific statement from the American Heart Association.”
48 Gerald F Fletcher et al., “Exercise standards for testing and training: a statement for healthcare professionals from the American Heart Association,” Circulation 104, no. 14 (2001).
49 Zuhl, “Tips for Monitoring Aerobic Exercise Intensity.”
50 Noortje Creemers et al., “The physiological mechanism behind the talk test,” Kinesiology 49, no. 1. (2017).
51 Rodríguez-Marroyo et al., “Relationship Between the Talk Test and Ventilatory Thresholds in Well-Trained Cyclists.”
52 Jaimie Ballweg et al., “Reliability of the talk test as a surrogate of ventilatory and respiratory compensation thresholds,” Journal of sports science & medicine 12, no. 3 (2013).
53 Carl Foster et al., “The talk test as a marker of exercise training intensity,” Journal of cardiopulmonary rehabilitation and prevention 28, no. 1 (2008).
54 Pedro T. Recalde, Foster, Carl, Skemp-Arlt, Karen M., Fater, Dennis CW, Neese, Christine A., Dodge, Christopher and John P Porcari, “The talk test as a simple marker of ventilatory threshold,” South African Journal of Sports Medicine 9, no. 2 (2002).
55 James EM Blackwell, “Exploring the efficacy of short-term high intensity interval training (HIIT) for improving surgical risk factors in older adults” (University of Nottingham, 2019).
56 Patrick Müller et al., “Lactate and BDNF: key mediators of exercise induced neuroplasticity?,” Journal of Clinical Medicine 9, no. 4 (2020).
57 Matevž Arčon and Nejc Šarabon, “A Minimalistic Approach to Promote Health-Span via Bouts of Daily Physical Activity in Older Adults: A Review,” Sport Mont 23, no. 2 (2025).
58 Zuhl, “Tips for Monitoring Aerobic Exercise Intensity.”
59 Raaj Kishore Biswas et al., “Wearable device-based health equivalence of different physical activity intensities against mortality, cardiometabolic disease, and cancer,” Nature Communications 16, no. 1 (2025/10/07 2025), https://doi.org/10.1038/s41467-025-63475-2, https://doi.org/10.1038/s41467-025-63475-2.
60 Oliver Bembom and Mark J van der Laan, “A practical illustration of the importance of realistic individualized treatment rules in causal inference,” Electronic journal of statistics 1 (2007).
61 Kasiakogias and Sharma, “Exercise: The ultimate treatment to all ailments?.”
62 Biswas et al., “Wearable device-based health equivalence of different physical activity intensities against mortality, cardiometabolic disease, and cancer.”
63 Minjae Yoon et al., “Association of physical activity level with risk of dementia in a nationwide cohort in Korea,” JAMA network open 4, no. 12 (2021).
64 Klaus Gebel et al., “Effect of moderate to vigorous physical activity on all-cause mortality in middle-aged and older Australians,” JAMA internal medicine 175, no. 6 (2015).
65 Biswas et al., “Wearable device-based health equivalence of different physical activity intensities against mortality, cardiometabolic disease, and cancer.”
66 “What Counts as Physical Activity for Adults,” Centers for Disease Control, Physical Activity Basics, US Centers for Disease Control, updated 2024/10/29/T16:21:16Z, 2023, https://www.cdc.gov/physical-activity-basics/adding-adults/what-counts.html.
67 “Physical activity,” World Health Organization, World Health Organization, https://www.who.int/initiatives/behealthy/physical-activity.
68 Biswas et al., “Wearable device-based health equivalence of different physical activity intensities against mortality, cardiometabolic disease, and cancer.”
69 Ulf Ekelund et al., “Dose–response associations, physical activity intensity and mortality risk: a narrative review,” Journal of sport and health science 13, no. 1 (2024).
70 Patrick Wahl, Wilhelm Bloch, and Sebastian Proschinger, “The molecular signature of high-intensity training in the human body,” International journal of sports medicine 43, no. 03 (2022).
71Khalilimeybodi et al., “Systems modeling of mitochondrial dynamics in different exercise regimes.”
72 Chen et al., “Kinase–Phosphatase Balance in Exercise Adaptation: Phosphorylation Programs, PTM Crosstalk, and Actionable Gaps.”
73 Biswas et al., “Wearable device-based health equivalence of different physical activity intensities against mortality, cardiometabolic disease, and cancer.”
74 Biswas et al., “Wearable device-based health equivalence of different physical activity intensities against mortality, cardiometabolic disease, and cancer.”
75 Courtney M Peterson, Darcy L Johannsen, and Eric Ravussin, “Skeletal muscle mitochondria and aging: a review,” Journal of aging research 2012, no. 1 (2012).
76 Arčon and Šarabon, “A Minimalistic Approach to Promote Health-Span via Bouts of Daily Physical Activity in Older Adults: A Review.”
77 Nichola J Conlon, “The role of NAD+ in regenerative medicine,” Plastic and Reconstructive Surgery 150 (2022).
78 J Sarah Crowley, Amy Liu, and Marek Dobke, “Regenerative and stem cell-based techniques for facial rejuvenation,” Experimental Biology and Medicine 246, no. 16 (2021).
79 Tom Benson et al., “Restoration of Complex Biological Components,” (2024).
80 David Andrew Gunn et al., “Mortality is Written on the Face,” The Journals of Gerontology: Series A 71, no. 1 (2015), https://doi.org/10.1093/gerona/glv090, https://doi.org/10.1093/gerona/glv090.
81 Minja Belic, Kevin Schneider, and David Furman, “Facial photographs as proxies for inflammatory aging,” bioRxiv (2025).
82 Jordan Bortz et al., “Biological age estimation using circulating blood biomarkers,” Communications Biology 6, no. 1 (2023/10/26 2023), https://doi.org/10.1038/s42003-023-05456-z, https://doi.org/10.1038/s42003-023-05456-z.
83 Zuyun Liu et al., “A new aging measure captures morbidity and mortality risk across diverse subpopulations from NHANES IV: a cohort study,” PLoS medicine 15, no. 12 (2018).
84 Gen Li et al., “Predicting healthspan and disease risks through biological age,” Trends in Molecular Medicine (2025).
85 Joshua J Levy et al., “Insights to aging prediction with AI based epigenetic clocks,” Epigenomics 17, no. 1 (2025).
86 Julia W Wu et al., “Biological age in healthy elderly predicts aging-related diseases including dementia,” Scientific reports 11, no. 1 (2021).
87 Kaare Christensen et al., “Perceived age as clinically useful biomarker of ageing: cohort study,” Bmj 339 (2009).
88 Jenna M Leser et al., “Aging, osteo-sarcopenia, and musculoskeletal mechano-transduction,” Frontiers in Rehabilitation Sciences 2 (2021).
89 Y Esquirol et al., “P1548 Forehead Wrinkles and risk of all-cause and cardiovascular mortality over 20-year follow-up in working population: VISAT study,” European Heart Journal 39, no. suppl_1 (2018).
90 Justin D. Crane et al., “Exercise-stimulated interleukin-15 is controlled by AMPK and regulates skin metabolism and aging,” Aging Cell 14, no. 4 (2015), https://doi.org/https://doi.org/10.1111/acel.12341, https://onlinelibrary.wiley.com/doi/abs/10.1111/acel.12341.
91 Crane et al., “Exercise-stimulated interleukin-15 is controlled by AMPK and regulates skin metabolism and aging.”
92 Noriaki Nakagawa et al., “The relationship between habitual physical activity and skin mechanical properties,” Skin Research and Technology 27, no. 3 (2021), https://doi.org/https://doi.org/10.1111/srt.12950, https://onlinelibrary.wiley.com/doi/abs/10.1111/srt.12950.
93 Hoffman et al., “Phosphoproteomics uncovers exercise intensity-specific skeletal muscle signaling networks underlying high-intensity interval training in healthy male participants.”
94 Blackwell, “Exploring the efficacy of short-term high intensity interval training (HIIT) for improving surgical risk factors in older adults.”
95 Torma et al., “High intensity interval training and molecular adaptive response of skeletal muscle.”
96 Gregory N Ruegsegger et al., “High-intensity aerobic, but not resistance or combined, exercise training improves both cardiometabolic health and skeletal muscle mitochondrial dynamics,” Journal of Applied Physiology 135, no. 4 (2023).
97 Khalilimeybodi et al., “Systems modeling of mitochondrial dynamics in different exercise regimes.”
98 Peterson, Johannsen, and Ravussin, “Skeletal muscle mitochondria and aging: a review.”
99 Pirani et al., “Beneficial mitochondrial biogenesis in gastrocnemius muscle promoted by high-intensity interval training in elderly female rats.”
100 Matthew M Robinson et al., “Enhanced protein translation underlies improved metabolic and physical adaptations to different exercise training modes in young and old humans,” Cell metabolism 25, no. 3 (2017).
101 Pirani et al., “Beneficial mitochondrial biogenesis in gastrocnemius muscle promoted by high-intensity interval training in elderly female rats.”
102 Yan Qiu et al., “Exercise sustains the hallmarks of health,” Journal of sport and health science 12, no. 1 (2023).
103 Yang Zhu et al., “Advances in exercise to alleviate sarcopenia in older adults by improving mitochondrial dysfunction,” Frontiers in Physiology 14 (2023).
104 Yuqing Li, Wanjun Zhao, and Qi Yang, “Effects of high-intensity interval training and moderate-intensity continuous training on mitochondrial dynamics in human skeletal muscle,” Frontiers in Physiology 16 (2025).
105 Philip M Batterson et al., “Two weeks of high-intensity interval training increases skeletal muscle mitochondrial respiration via complex-specific remodeling in sedentary humans,” Journal of applied physiology (2023).
106 Carlos López-Otín et al., “The hallmarks of aging,” Cell 153, no. 6 (2013).
107 Bennett G Childs et al., “Senescent cells: an emerging target for diseases of ageing,” Nature reviews Drug discovery 16, no. 10 (2017).
108 Bente K Pedersen and Mark A Febbraio, “Muscle as an endocrine organ: focus on muscle-derived interleukin-6,” Physiological reviews (2008).
109 Rosa Mancinelli et al., “Biological aspects of selected myokines in skeletal muscle: focus on aging,” International journal of molecular sciences 22, no. 16 (2021).
110 Weiping Xiao et al., “Exercise-induced irisin ameliorates cognitive impairment following chronic cerebral hypoperfusion by suppressing neuroinflammation and hippocampal neuronal apoptosis,” Journal of Neuroinflammation 22, no. 1 (2025).
111 Graziana Colaianni et al., “Irisin correlates positively with BMD in a cohort of older adult patients and downregulates the senescent marker p21 in osteoblasts,” Journal of Bone and Mineral Research 36, no. 2 (2020).
112 Cailin Wang et al., “Irisin inhibits microglial senescence via TFAM-mediated mitochondrial metabolism in a mouse model of tauopathy,” Immunity & Ageing 21, no. 1 (2024).
113 Atefe Torabi et al., “Differences in the impact of various types of exercise on irisin levels: a systematic review and meta-analysis,” International Journal of Preventive Medicine 15 (2024).
114 Noni T Frankenberg et al., “Exercise attenuates stress-related signaling as sensed by higher phosphorylation of small heat shock proteins in skeletal muscle from older individuals,” Journal of Sport and Health Science (2025).
115 Frankenberg et al., “Exercise attenuates stress-related signaling as sensed by higher phosphorylation of small heat shock proteins in skeletal muscle from older individuals.”
116 Amal A Wanigatunga et al., “Moderate-to-Vigorous physical activity at any dose reduces all-cause dementia risk regardless of frailty status,” Journal of the American Medical Directors Association 26, no. 3 (2025).
117 Mitzi M Gonzales et al., “Biological aging processes underlying cognitive decline and neurodegenerative disease,” The Journal of clinical investigation 132, no. 10 (2022).
118 Joshua T Reycraft et al., “Exercise intensity and recovery on circulating brain-derived neurotrophic factor,” Med Sci Sports Exerc 52, no. 5 (2020).
119 Kaihang Liu et al., “The effects of high-intensity interval training on cognitive performance: a systematic review and meta-analysis,” Scientific Reports 14, no. 1 (2024).
120 Gonzales et al., “Biological aging processes underlying cognitive decline and neurodegenerative disease.”
121 Pierre J Magistretti and Igor Allaman, “Lactate in the brain: from metabolic end-product to signalling molecule,” Nature reviews neuroscience 19, no. 4 (2018).
122 Xiangli Xue et al., “The potential mechanisms of lactate in mediating exercise-enhanced cognitive function: a dual role as an energy supply substrate and a signaling molecule,” Nutrition & metabolism 19, no. 1 (2022).
123 Gonzales et al., “Biological aging processes underlying cognitive decline and neurodegenerative disease.”
124 “Brain changes seen in cabbies who take ‘The Knowledge’,” BBC News, BBC, updated 2011/12/08/, 2011, https://www.bbc.com/news/health-16086233.
125 “Cache Cab: Taxi Drivers’ Brains Grow to Navigate London’s Streets,” Ferris Jabr, Scientific American, 2026, https://www.scientificamerican.com/article/london-taxi-memory/.
126 Eleanor A. Maguire et al., “Navigation-related structural change in the hippocampi of taxi drivers,” Proceedings of the National Academy of Sciences 97, no. 8 (2000), https://doi.org/doi:10.1073/pnas.070039597, https://www.pnas.org/doi/abs/10.1073/pnas.070039597.
127 Rossana Gómez-Campos et al., “Relationship between age and handgrip strength: Proposal of reference values from infancy to senescence,” Frontiers in public health 10 (2023).
128 Darryl P Leong et al., “Prognostic value of grip strength: findings from the Prospective Urban Rural Epidemiology (PURE) study,” The lancet 386, no. 9990 (2015).
129 Bente K Pedersen and Mark A Febbraio, “Muscles, exercise and obesity: skeletal muscle as a secretory organ,” Nature Reviews Endocrinology 8, no. 8 (2012).
130 Anne Marie W Petersen and Bente Klarlund Pedersen, “The anti-inflammatory effect of exercise,” Journal of applied physiology 98, no. 4 (2005).
131 Colaianni et al., “Irisin correlates positively with BMD in a cohort of older adult patients and downregulates the senescent marker p21 in osteoblasts.”
132 Laura A Schaap et al., “Inflammatory markers and loss of muscle mass (sarcopenia) and strength,” The American journal of medicine 119, no. 6 (2006).
133 Mark D. Peterson et al., “Grip strength is inversely associated with DNA methylation age acceleration,” Journal of Cachexia, Sarcopenia and Muscle 14, no. 1 (2023), https://doi.org/https://doi.org/10.1002/jcsm.13110, https://onlinelibrary.wiley.com/doi/abs/10.1002/jcsm.13110.
134 Mark D. Peterson et al., “Resistance exercise for muscular strength in older adults: A meta-analysis,” Ageing Research Reviews 9, no. 3 (2010/07/01/ 2010), https://doi.org/https://doi.org/10.1016/j.arr.2010.03.004, https://www.sciencedirect.com/science/article/pii/S1568163710000279.
135 Nina T. Rogers et al., “Physical activity and trajectories of frailty among older adults: Evidence from the English Longitudinal Study of Ageing,” PLOS ONE 12, no. 2 (2017), https://doi.org/10.1371/journal.pone.0170878, https://doi.org/10.1371/journal.pone.0170878.
136 C. H. Turner, “Three rules for bone adaptation to mechanical stimuli,” Bone 23, no. 5 (1998/11/01/ 1998), https://doi.org/https://doi.org/10.1016/S8756-3282(98)00118-5, https://www.sciencedirect.com/science/article/pii/S8756328298001185.
137 “Meta-analysis: Excess Mortality After Hip Fracture Among Older Women and Men,” Annals of Internal Medicine 152, no. 6 (2010), https://doi.org/10.7326/0003-4819-152-6-201003160-00008 %m 20231569, https://www.acpjournals.org/doi/abs/10.7326/0003-4819-152-6-201003160-00008.
138 Daniel Pincus et al., “Association Between Wait Time and 30-Day Mortality in Adults Undergoing Hip Fracture Surgery,” JAMA 318, no. 20 (2017), https://doi.org/10.1001/jama.2017.17606, https://doi.org/10.1001/jama.2017.17606.
139 Gregory M. Fahy et al., “Reversal of epigenetic aging and immunosenescent trends in humans,” Aging Cell 18, no. 6 (2019), https://doi.org/https://doi.org/10.1111/acel.13028, https://onlinelibrary.wiley.com/doi/abs/10.1111/acel.13028.
140 Chris Curwen, “Dr. Greg Fahy on Reversing Human Aging | Our Longevity Futures, with Chris Curwen | Ep.2,” in Gowinglife (YouTube, 2021). https://www.youtube.com/watch?v=rpwtQ-DG2tE.
141 Donald B. Palmer, “The Effect of Age on Thymic Function,” Mini Review, Frontiers in Immunology Volume 4 – 2013 (2013-October-07 2013), https://doi.org/10.3389/fimmu.2013.00316, https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2013.00316.
142 J. Gui et al., “Thymus Size and Age-related Thymic Involution: Early Programming, Sexual Dimorphism, Progenitors and Stroma,” Aging Dis 3, no. 3 (Jun 2012).
143 Paula J. Busse and Sameer K. Mathur, “Age-related changes in immune function: Effect on airway inflammation,” Journal of Allergy and Clinical Immunology 126, no. 4 (2010/10/01/ 2010), https://doi.org/https://doi.org/10.1016/j.jaci.2010.08.011, https://www.sciencedirect.com/science/article/pii/S0091674910011966.
144 Beata Berent-Maoz, Encarnacion Montecino-Rodriguez, and Kenneth Dorshkind, “Genetic regulation of thymocyte progenitor aging,” Seminars in Immunology 24, no. 5 (2012/10/01/ 2012), https://doi.org/https://doi.org/10.1016/j.smim.2012.04.006, https://www.sciencedirect.com/science/article/pii/S1044532312000450.
145 Busse and Mathur, “Age-related changes in immune function: Effect on airway inflammation.”
146 Sam Palmer et al., “Thymic involution and rising disease incidence with age,” Proceedings of the National Academy of Sciences 115, no. 8 (2018), https://doi.org/doi:10.1073/pnas.1714478115, https://www.pnas.org/doi/abs/10.1073/pnas.1714478115.
147 Palmer et al., “Thymic involution and rising disease incidence with age.”
148 Fahy et al., “Reversal of epigenetic aging and immunosenescent trends in humans.”
149 Fahy et al., “Reversal of epigenetic aging and immunosenescent trends in humans.”
150 Inc Intervene Immune, Thymus Regeneration, Immunorestoration, and Insulin Mitigation Extension Trial, clinicaltrials.gov (2025/05/05/ 2025), https://clinicaltrials.gov/study/NCT04375657.
151 “Reversing the aging pocess with epigenetics?,” BASF, BASF, 2025, https://www.basf.com/global/en/media/magazine/creatingchemistrystories/2025/stopping-the-clock.
152 Fahy et al., “Reversal of epigenetic aging and immunosenescent trends in humans.”
153 Richard J. Godfrey, Zahra Madgwick, and Gregory P. Whyte, “The Exercise-Induced Growth Hormone Response in Athletes,” Sports Medicine 33, no. 8 (2003/07/01 2003), https://doi.org/10.2165/00007256-200333080-00005, https://doi.org/10.2165/00007256-200333080-00005.
154 Cathy J. Pritzlaff et al., “Impact of acute exercise intensity on pulsatile growth hormone release in men,” Journal of Applied Physiology 87, no. 2 (1999), https://doi.org/10.1152/jappl.1999.87.2.498, https://journals.physiology.org/doi/abs/10.1152/jappl.1999.87.2.498.
155 Laurie Wideman et al., “Growth Hormone Release During Acute and Chronic Aerobic and Resistance Exercise,” Sports Medicine 32, no. 15 (2002/12/01 2002), https://doi.org/10.2165/00007256-200232150-00003, https://doi.org/10.2165/00007256-200232150-00003.
156 Dennis D. Taub, William J. Murphy, and Dan L. Longo, “Rejuvenation of the aging thymus: growth hormone-mediated and ghrelin-mediated signaling pathways,” Current Opinion in Pharmacology 10, no. 4 (2010/08/01/ 2010), https://doi.org/https://doi.org/10.1016/j.coph.2010.04.015, https://www.sciencedirect.com/science/article/pii/S1471489210000512.
157 Todd M. Manini et al., “Growth hormone responses to acute resistance exercise with vascular restriction in young and old men,” Growth Hormone & IGF Research 22, no. 5 (2012/10/01/ 2012), https://doi.org/https://doi.org/10.1016/j.ghir.2012.05.002, https://www.sciencedirect.com/science/article/pii/S1096637412000470.
158 Niharika Arora Duggal et al., “Major features of immunesenescence, including reduced thymic output, are ameliorated by high levels of physical activity in adulthood,” Aging Cell 17, no. 2 (2018), https://doi.org/https://doi.org/10.1111/acel.12750, https://onlinelibrary.wiley.com/doi/abs/10.1111/acel.12750.
159 Zsolt Radak et al., “Exercise, oxidants, and antioxidants change the shape of the bell-shaped hormesis curve,” Redox Biology 12 (2017/08/01/ 2017), https://doi.org/https://doi.org/10.1016/j.redox.2017.02.015, https://www.sciencedirect.com/science/article/pii/S2213231716304700.
160 Zhengbiao Xue et al., “Dose-response relationship of cardiorespiratory fitness with incident atrial fibrillation,” Heart Failure Reviews 25, no. 3 (2020/05/01 2020), https://doi.org/10.1007/s10741-019-09871-5, https://doi.org/10.1007/s10741-019-09871-5.
161 Jonas Zacher et al., “Exercise and Atrial Fibrillation: The Dose Makes the Poison? A Narrative Review,” Int J Sports Med 45, no. 01 (2023/10/06 2023), https://doi.org/10.1055/a-2152-7628, http://www.thieme-connect.com/products/ejournals/abstract/10.1055/a-2152-7628.
162 William Newman et al., “Risk of atrial fibrillation in athletes: a systematic review and meta-analysis,” British Journal of Sports Medicine 55, no. 21 (2021), https://doi.org/10.1136/bjsports-2021-103994, https://bjsm.bmj.com/content/bjsports/55/21/1233.full.pdf.
163 Naiara Calvo et al., “Emerging risk factors and the dose–response relationship between physical activity and lone atrial fibrillation: a prospective case–control study,” EP Europace 18, no. 1 (2016), https://doi.org/10.1093/europace/euv216, https://doi.org/10.1093/europace/euv216.
164 Newman et al., “Risk of atrial fibrillation in athletes: a systematic review and meta-analysis.”
165 O. A. Centurión et al., “The Association Between Atrial Fibrillation and Endurance Physical Activity: How Much is too Much?,” J Atr Fibrillation 12, no. 3 (Oct-Nov 2019), https://doi.org/10.4022/jafib.2167.
166 Zacher et al., “Exercise and Atrial Fibrillation: The Dose Makes the Poison? A Narrative Review.”
167 Calvo et al., “Emerging risk factors and the dose–response relationship between physical activity and lone atrial fibrillation: a prospective case–control study.”
168 Jeffrey B. Kreher and Jennifer B. Schwartz, “Overtraining Syndrome:A Practical Guide,” Sports Health 4, no. 2 (2012), https://doi.org/10.1177/1941738111434406, https://journals.sagepub.com/doi/abs/10.1177/1941738111434406.
169 A. Angeli et al., “The overtraining syndrome in athletes: A stress-related disorder,” Journal of Endocrinological Investigation 27, no. 6 (2004/06/01 2004), https://doi.org/10.1007/BF03347487, https://doi.org/10.1007/BF03347487.
170 Angeli et al., “The overtraining syndrome in athletes: A stress-related disorder.”
171 Flavio A. Cadegiani and Claudio E. Kater, “Hypothalamic-Pituitary-Adrenal (HPA) Axis Functioning in Overtraining Syndrome: Findings from Endocrine and Metabolic Responses on Overtraining Syndrome (EROS)—EROS-HPA Axis,” Sports Medicine – Open 3, no. 1 (2017/12/08 2017), https://doi.org/10.1186/s40798-017-0113-0, https://doi.org/10.1186/s40798-017-0113-0.
172 Sergio Gutierrez Nunez et al., “Chronic Stress and Autoimmunity: The Role of HPA Axis and Cortisol Dysregulation,” International Journal of Molecular Sciences 26, no. 20 (2025), https://www.mdpi.com/1422-0067/26/20/9994.
173 Thomas L. Stöggl and Billy Sperlich, “The training intensity distribution among well-trained and elite endurance athletes,” Focused Review, Frontiers in Physiology Volume 6 – 2015 (2015-October-27 2015), https://doi.org/10.3389/fphys.2015.00295, https://www.frontiersin.org/journals/physiology/articles/10.3389/fphys.2015.00295.
174 Stephen Seiler, “What is Best Practice for Training Intensity and Duration Distribution in Endurance Athletes?,” International Journal of Sports Physiology and Performance 5, no. 3 (01 Sep. 2010 2010), https://doi.org/10.1123/ijspp.5.3.276, https://journals.humankinetics.com/view/journals/ijspp/5/3/article-p276.xml.
175 Thomas Stöggl and Billy Sperlich, “Polarized training has greater impact on key endurance variables than threshold, high intensity, or high volume training,” Original Research, Frontiers in Physiology Volume 5 – 2014 (2014-February-04 2014), https://doi.org/10.3389/fphys.2014.00033, https://www.frontiersin.org/journals/physiology/articles/10.3389/fphys.2014.00033.
176 Iker Muñoz et al., “Does Polarized Training Improve Performance in Recreational Runners?,” International Journal of Sports Physiology and Performance 9, no. 2 (01 Mar. 2014 2014), https://doi.org/10.1123/ijspp.2012-0350, https://journals.humankinetics.com/view/journals/ijspp/9/2/article-p265.xml.
177 Fiona C Bull et al., “World Health Organization 2020 guidelines on physical activity and sedentary behaviour,” British Journal of Sports Medicine 54, no. 24 (2020), https://doi.org/10.1136/bjsports-2020-102955, https://bjsm.bmj.com/content/bjsports/54/24/1451.full.pdf.
178 Richard D Olson et al., “Physical activity guidelines for Americans,” REVISTA NADAR! SWIMMING MAGAZINE-Periódico científico em esportes e fitness aquático-natação, pólo aquático, nado sincronizado, saltos ornamentais, travessias aquáticas 3 (2023).
179 Martin J Gibala et al., “Physiological adaptations to low-volume, high-intensity interval training in health and disease,” The Journal of physiology 590, no. 5 (2012).
180 MacInnis and Gibala, “Physiological adaptations to interval training and the role of exercise intensity.”





